|
Michio HASHIMOTO
[Abstract]
|
|
Docosahexaenoic acid (DHA, 22:6n-3) is one of the n-3 polyunsaturated
fatty acids (n-3 PUFA) found plentifully in lipids contained in marine
products such as fish oils. Since it was reported that the prevalence of
myocardial infarction among the primarily fish-eating Greenland natives
was much lower than that among primarily meat-eating westerners, n-3 PUFA
have been vigorously studied. In addition to DHA, eicosapentaenoic acid
is another well-known example of n-3 PUFA. Currently, many reports on the
former deal with the central nervous system (CNS), while many on the latter
are concerned with the circulatory system.
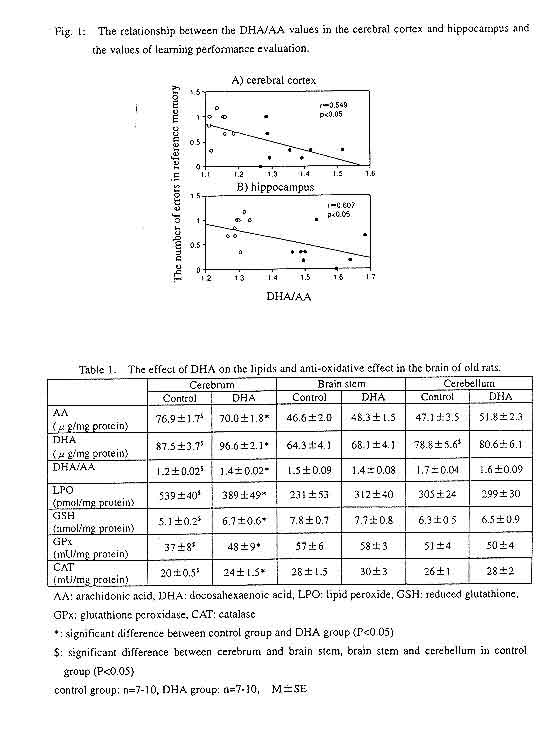
Among brain lipids, the main n-6 PUFA is arachidonic acid, and the main n-3 PUFA is DHA. The sum of these two fatty acids accounts for 50% of the total fatty acid amount in the cerebral cortex, although the figure is different for different portions of the brain. The relationship between DHA and hyperfunctions of the brain has thus attracted attention. Studies on people include many experiments involving premature infants, one of which reported that a group of premature infants raised with DHA-enhanced mother's milk or infant formula later developed a higher IQ and better vision than a group not receiving DHA-enhanced milk, suggesting an advantage of DHA intake1). Another study showed that the DHA content in the hippocampus of patients who died of Alzheimer's dementia was remarkably lower than that of patients who died of other types of dementia2). The study also suggested an association between the decreased DHA level in the brain and Alzheimer's dementia. With regard to animal experiments, many researchers have already shown that a diet deficient in n-3 fatty acids causes deterioration of memory as well as decreased DHA levels in the blood and brain1),3). Although details of these effects of DHA on the CNS remain to be elucidated, recently it has been reported that in rats deficient in n-3 fatty acids, the density of synaptic vesicles in the terminals of the hippocampus CA1 region decreased4), an intriguing finding in terms of investigating the functional mechanism of DHA on the CNS. However, while many of these studies investigated the effect of DHA (or n-3 PUFA) deficiency, there are few which examined the effect of DHA intake. Recently, we have investigated the effect of DHA intake on brain functions using both young and old rats. First, in order to examine the effect of DHA on the memory of rats, a maze experiment was conducted using both young (5 weeks old) and old (100 weeks old) male Wistar rats in the third generation of the rats maintained through three generations with food not containing fish oil5). DHA was orally administered to the animals (300 mg/kg/day). The maze had eight radiating arms. At the end of four randomly chosen arms a dietary pellet was placed in a hole. The behavior of the rats was then observed until they had taken all the pellets in the maze. The animals' learning performance was determined in terms of the following three values: the number of correct choices (the number of times a rat entered an arm with a pellet within the first four choices), the number of errors in reference memory (the number of times a rat entered an arm without a pellet), and the number of errors in working memory (the number of times a rat re-entered an arm which it had entered previously). The results showed a significantly greater number of correct choices and a smaller number of errors in reference memory in the DHA-fed young group than in the DHA-deficient young group. Similarly, in the older groups, the DHA-fed group showed a greater number of correct choices and a smaller number of errors in reference memory than the DHA-deficient group. In addition, the DHA-fed older group showed a smaller number of errors in working memory. A comparison between the young and old rats revealed that aging caused a decreased number of correct choices and significantly greater numbers of errors in both reference and working memories. The number of errors in reference memory decreases if a rat remembers the arms containing rewards until the next session. Therefore, reference memory performance was established by evaluating the number of correct choices and that of errors in reference memory. On the other hand, the number of errors in working memory decreases if a rat remembers the arms it has entered before. Thus working memory performance was established by evaluating the number of errors in working memory. Our study results showed smaller numbers of errors in both reference and working memories in the DHA-fed group than in the DHA- deficient group, suggesting that the administration of DHA improved both reference and working memories. Although these memories declined from aging, the administration of DHA improved them in the older group as well as in the young group, suggesting that this effect of DHA is not affected by aging. It is thus expected that DHA might have an effect in improving senile dementia. After the evaluation of memory, the fatty acid levels in the blood serum and brain of the young rats were determined in order to examine the relation between fatty acid levels and performance on the maze task5). The DHA level in the blood serum of the DHA-fed group was more than double that of the DHA-deficient group, while the arachidonic acid (AA) level in the former was lower by approximately 45% than that of the latter. In the brain, while the DHA levels in the cerebral cortex and hippocampus were significantly higher in the DHA-fed group, there was no significant difference in the DHA levels in the brain stem and cerebellum. Despite the lower level of AA in the blood serum of the DHA-fed group, no difference was found in the AA levels in any part of the brain between the two groups. However, the ratio of DHA to AA level (DHA/AA) was significantly greater in both the cerebral cortex and hippocampus of the DHA-fed group. The examination of the relation between the fatty acid levels in the brain and the performance on the maze task revealed a significant negative correlation between the number of errors in reference memory and the DHA/AA values in both the cerebral cortex and hippocampus (Fig. 1). As these two are the portions of the brain where memory accumulates, it can be inferred that the increased levels of DHA improved memory by playing a part in the maturation or promotion of the functions of these portions of the brain. It is inferred that PUFA are oxidized in the body by reactive oxygen species and free radicals, which are then amplified by radical chain reactions through lipid peroxides, causing damage in cells. Therefore, we then examined the effect of DHA intake on the peroxidization of lipids in the brain. DHA was orally administered to old (100 weeks old) Wistar rats (300 mg/kg/day), and the levels of fatty acid, lipid peroxides (LPO), reduced glutathione (GSH), and two anti-oxidative enzymes (catalase (CAT) and glutathione peroxidase (GPx)) in the cerebrum, cerebellum, and brain stem were determined 12 weeks after the administration6),7). In a DHA-deficient group, the LPO level in the cerebrum was higher than in the cerebellum and brain stem, and the levels of the anti-oxidative agents, namely, GSH, CAT and GPx, were the lowest in the cerebrum. In the DHA-fed group, while the DHA level in the cerebrum was significantly higher than that of the DHA-deficient group, a lower level of LPO and higher levels of GSH, CAT and GPx in the cerebrum were found (Table 1). Moreover, in the cerebrum, a significant negative correlation was found between the LPO level and DHA/AA value (r=-0.792, p<0.05), whereas a significant positive correlation was shown between the levels of GPx and CAT and the DHA/AA value (r=0.730, p<0.05; r=0.870, p<0.05). It can be inferred from these results that DHA promotes anti-oxidation in the brain and inhibits the production of LPO in the cerebrum. PUFA are susceptible to oxidation when exposed to air, and in the body they might be peroxidized by such agents as reactive oxygen species. Thus, it has been pointed out that accumulated lipid peroxides might cause damage in cells, constituting a potential risk factor of geriatric disorders including arteriosclerosis. However, our results suggested a possibility that, instead of being peroxidized, DHA in the brain serves as a radical scavenger which promotes anti-oxidation and protects nerve cells from harmful effects including oxidation stress caused by reactive oxygen species, thus playing a part in the formation and maintenance of memory. Recently, Yavine et al. found that the production of lipid peroxides in the fetal rat brain was inhibited by the administration of DHA through amniocentesis, suggesting that DHA might have an effect in protecting against oxidation stress occurring after the cerebral ischemia and subsequent resumption of perfusion experienced by the fetal brain during delivery8). As the DHA content in the brain is much higher than that in other organs, the elucidation of the physiological functions of DHA in the brain will become an increasingly intriguing subject in terms of understanding the complicated functions of the brain. |
|
Toward a new recommended dietary allowance for vitamin
C based on antioxidant and health effects in humans
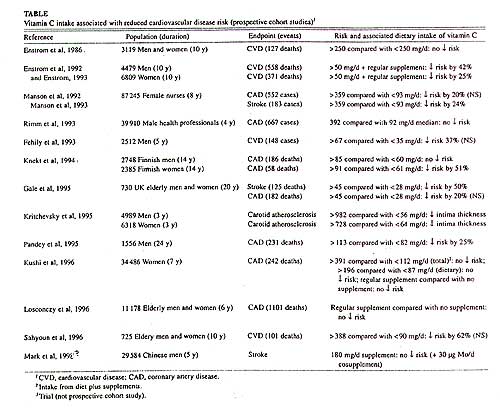
The current recommended dietary allowance (RDA) for vitamin C for adult nonsmoking men and women is 60 mg/d, which is based on a mean requirement of 46 mg/d to prevent the deficiency disease scurvy. However, recent scientific evidence indicates that an increased intake of vitamin C is associated with a reduced risk of chronic diseases such as cancer, cardiovascular disease, and cataract, probably through antioxidant mechanisms. It is likely that the amount of vitamin C required to prevent scurvy is not sufficient to optimally protect against these diseases. Because the RDA is defined as "the average daily dietary intake level that is sufficient to meet the nutrient requirement of nearly all healthy individuals in a group," it is appropriate to reevaluate the RDA for vitamin C. Therefore, we reviewed the biochemical, clinical, and epidemiologic evidence to date for a role of vitamin C in chronic disease prevention. Over the past 15y, several prospective cohort studies have been published on the association between vitamin C intake and the risk of cardiovascular disease (Table). The totality of the reviewed data suggests that an intake of 90-100 mg vitamin C/d is required for optimum reduction of chronic disease risk in nonsmoking men and women. This amount is about twice on which the current RDA for vitamin C is based, suggesting a new RDA of 120 mg vitamin C/d. |
 |
|
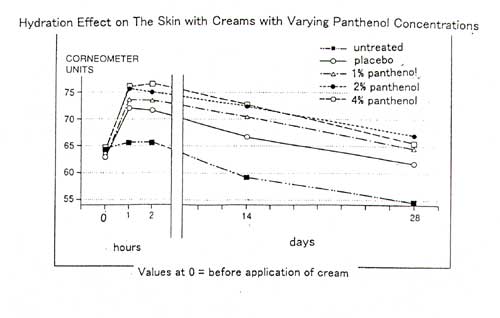
excerpt from Report on the Efficacy of Various Cream Formulations Containing: PANTHENOL (commissioned by F. Hoffmann-La Roche Ltd)
The study was double-blind and randomized. The preparations tested were
creams containing 1%, 2%, or 4% panthenol, and a placebo cream. The test
persons applied the creams twice daily as instructed on separate test areas
on the forearms. A control area remained untreated for the duration of
the study. Hydration of the stratum corneum was determined using the condensor
method. Measurements were made with a corneometer.
|
 |
|
from Nutritional Concerns of Women Oral contraceptives (OCs) are among the most thoroughly studied drugs.
In our modern society, women are becoming sexually active earlier, delaying
the birth of their first child and limiting their total number of children.
As a consequence, approximately 14 million women in the U.S. and 150 million
worldwide, averaging 85% of reproductive age women, choose oral contraceptives
for birth control because of their convenience and efficiency. The new
generation of OCs includes both the low-dose combined estrogen-progestogen
pill (containing less than 50μg estrogen and less than 1.5 mg progestogen)
as well as the progestogen-only pill. In the first category, more than
two dozen preparation are currently marketed. The majority are modifications
of the "first generation" higher dose combined estrogen-progestogen.
OC-Induced Metabolic Changes
|
 |